What are proteasomes ?
Proteasomes are cytosolic multisubunit proteases which are involved in
cell cycle control, transcription factor activation and the generation
of peptide ligands for MHC I molecules (for reviews, see Baumeister et
al. (1998), Rock & Goldberg (1999), Uebel & Tampe (1999)). They exist
in several forms; either as proteolytically active core complexes or 20S
proteasomes and, when associated with the ATP-dependent 19S cap
complexes, larger 26S proteasomes that are able to recognize
proteins marked by ubiquitin for proteasomal degradation (Jentsch &
Schlenker, 1995; Hershko & Ciechanover, 1998). Another protein complex
known to associate with the 20S core particle is PA28, the 11S
regulator (Ahn et al., 1995), which was shown to improve the yield of
antigenic peptides (Groettrup et al., 1996; Dick et al., 1996).
Eukaryotic 20S proteasomes consist of four stacked rings (overall
stoichiometry alpha7beta7beta7alpha7), each consisting of 7 different subunits
(Groll et al., 1997 [See picture taken from this reference at
the bottom of the page. The picture shows a section through the
cylinder of yeast
20S proteasomes. The positions of the active sites are highlighted
through binding of specific inhibitors (yellow).]) . Each of the two inner beta-rings carries three
catalytically active sites on its inner surface. Their proteolytic
specificities have been described as chymotrypsin-like (cleaving after
large, hydrophobic AAs), trypsin-like (cleaving after basic AAs) and
peptidyl-glutamyl-peptide-hydrolyzing (cleaving after acidic AAs) (for
review, see Uebel & Tampe (1999)). Strings of unfolded proteins are
thought to be inserted into the cylinder and to be cut into pieces by
the active sites; the resulting peptide fragments are then released
into the cytosol. Functionally, proteasomal protein degradation is
believed to proceed from one substrate end to the other
("processively"), without the release of large degradation
intermediates (Akopian et al., 1997; Nussbaum et al., 1998; Kisselev
et al., 1999).
Why is proteasomal cleavage specificity important for immune
responses?
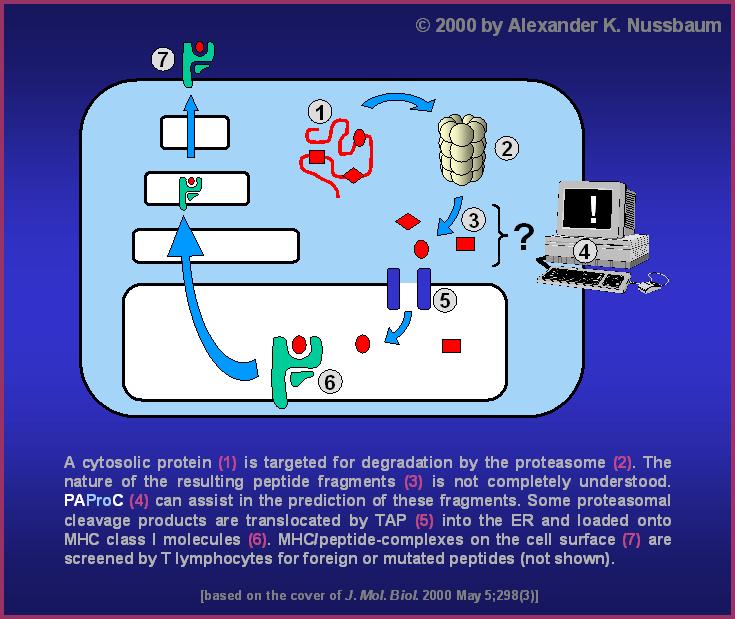
In vertebrate cells, some of the proteolytic fragments produced by
proteasomes are fed into the antigen processing machinery (see
picture ). Since
peptide presentation by MHC I molecules at the cell surface is an
intrinsic requirement for the ability of the immune system to
eradicate virus-infected or transformed cells (Rammensee et al., 1993;
Pamer & Cresswell, 1998), it is of general interest to know exactly
how the proteasome is involved in this process. Proteasomal cleavage
specificity has been assessed by in vitro digestion experiments using
either tri- or tetrapeptides with fluorogenic leaving groups
(Kuckelkorn et al., 1995; Heinemeyer et al., 1997; Arendt &
Hochstrasser, 1997), peptides of 15-40 AAs (Boes et al., 1994;
Niedermann et al., 1995; Niedermann et al., 1996; Dick et al., 1998),
or denatured proteins (Dick et al., 1991; Dick et al., 1994; Kisselev
et al., 1998, Kisselev et al., 1999) as substrates. We analyzed the
cleavage preferences of yeast wild-type and mutant proteasomes in a
non-modified protein (Nussbaum et al., 1998). Using statistical
analysis of cut sites, it was possible for the first time to determine
so-called cleavage motifs, i.e. the preferred sequences around
cleavage sites, for the three active beta-subunits of yeast proteasomes.
Why would a prediction tool be beneficial?
In order to apply experimentally determined information on cleavage
site selection by proteasomes to any possible proteasome substrate,
one needs an automated prediction device. Such devices already exist
for the binding of peptides to MHC I molecules (Database
SYFPEITHI ,
Rammensee et al., 1997) and have been described for peptide transport
by the transporter associated with antigen processing (TAP) (Daniel et
al., 1998). However, devices for the prediction of proteasomal
cleavages are only at the beginning of their development. A
proteasomal cleavage prediction tool could, especially in combination
with MHC ligand predictors as SYFPEITHI, help to improve the forecast
of MHC class I restricted CTL-responses. More specifically, it could
support researchers in their quest for individual CTL-epitopes by
limiting the number of possible MHC class I ligands from protein
antigens. In addition, the effect of amino acid mutations in viral or
tumor-specific proteins on antigen presentation could be
assessed. Thus, proteasomal cleavage prediction would lend a hand in
rational vaccine design.
PAProC
We have made the first step towards this end by providing PAProC
(Prediction Algorithm for Proteasomal Cleavages), a public prediction
tool for proteasomal cleavages. PAProC offers information on both the
general cleavability of amino acid sequences (cuts per amino acids)
and individual cleavages (positions and estimated strength; for
details, please refer to the
user information).
PAProC was developed from the beginning, i.e. from the experimental
basis to the ready-to-use public prediction tool, by proteasome
experts at the Department of Immunology in close collaboration with
programmers at the Department of Biomathematics, both at the
University of Tübingen, Germany. We are therefore confident that
PAProC has profited from the best possible expertise. However, we are
aware of the fact that PAProC is still in its teething stage. For
example, cleavage sites and estimated cleavage strength are not yet
based on quantified cleavage data (in PAProC I). Therefore, we are continuously
working to improve PAProC. However, we need your help: The program
will profit from your experience with it. So please let us know how
PAProC performed for you. Thank you for your collaboration.
Links
On our
link list you can find several pages concerning proteasome and more.
Last update: 13.4.2005
More detailed information about this program can be found in the
following publications:
C. Kuttler, A.K.
Nussbaum, T.P. Dick, H.-G. Rammensee, H. Schild, K.P. Hadeler, An algorithm
for the prediction of proteasomal cleavages,
J. Mol. Biol. 298 (2000), 417-429 ,
and
A.K. Nussbaum, C. Kuttler, K.P. Hadeler, H.-G. Rammensee, H. Schild, PAProC: A Prediction Algorithm for Proteasomal Cleavages available on the
WWW, Immunogenetics 53 (2001), 87-94
For comprehensive background information, please refer to
From the test tube to the World Wide Web - The cleavage specificity of
the proteasome (A.K. Nussbaum, dissertation, University of
Tuebingen, Germany, 2001).
The use of PAProC is restricted to non-commercial
purposes.

![Inner surface of the yeast 20S proteasome; yellow:
inhibitors bound to the three active sites [Groll et al. (1997), Nature].](http://www.paproc2.de/paproc1/20szit.jpg)